
 数据结构
数据结构 网络
网络 关系数据库管理系统 (RDBMS)
关系数据库管理系统 (RDBMS) 操作系统
操作系统 Java
Java iOS
iOS HTML
HTML CSS
CSS Android
Android Python
Python C 编程
C 编程 C++
C++ C#
C# MongoDB
MongoDB MySQL
MySQL Javascript
Javascript PHP
PHPII 类 MHC - 结构、合成及意义
简介
主要组织相容性复合体 (MHC) 是一组基因,编码参与将抗原呈递给免疫系统的细胞表面蛋白。这些蛋白分为两类:I 类 MHC 和 II 类 MHC。
II 类 MHC 分子主要存在于抗原呈递细胞 (APC) 上,例如树突状细胞、巨噬细胞和 B 细胞。它们通过将外源性抗原呈递给 CD4+ T 细胞,在启动和调节适应性免疫反应中发挥关键作用。
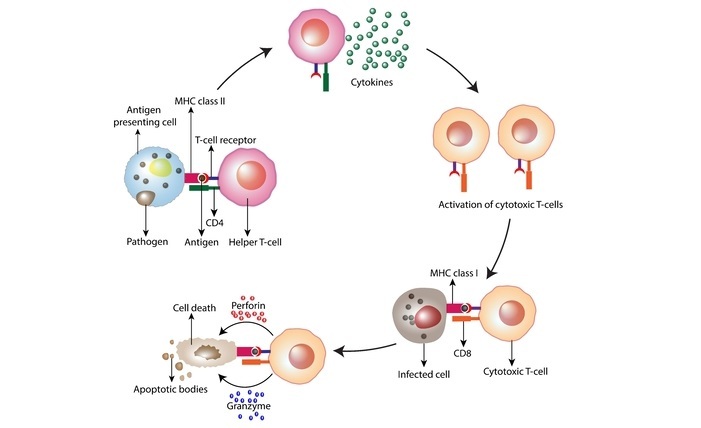
II 类 MHC 分子的结构
II 类 MHC 分子是由两条链组成的异二聚体:α 链和 β 链。α 链由 HLA-D 基因中的一个编码,而 β 链由 HLA-DP、HLA-DQ 或 HLA-DR 基因中的一个编码。每条链有两个结构域:细胞外结构域 (ECD) 和跨膜结构域 (TMD)。两条链的 TMD 都锚定在细胞的质膜上。
α 链的细胞外结构域由两个免疫球蛋白 (Ig) 样结构域组成,称为 α1 结构域和 α2 结构域。α1 结构域参与与 β 链的结合,对 II 类 MHC 分子的稳定性至关重要。
α2 结构域形成肽结合槽的底部,并负责与肽的大部分相互作用。β 链的细胞外结构域由一个免疫球蛋白样结构域(称为 β1 结构域)和一个 MHC II 特异性结构域(称为“β2 结构域”)组成。
β1 结构域参与与 α 链的结合,而 β2 结构域形成肽结合槽的顶部,并负责与肽的相互作用。II 类 MHC 分子的肽结合槽可以容纳长度为 13-25 个氨基酸的肽。该槽内衬有针对某些氨基酸的特定氨基酸残基,使肽结合槽能够结合多种肽。
肽结合槽的两端都是封闭的,防止肽脱落。α 链和 β 链之间的相互作用对 II 类 MHC 分子的稳定性至关重要。α1 和 β1 结构域以“凸块插入孔”的方式相互作用,而 β2 结构域通过氢键和盐桥网络与 α2 结构域相互作用。
α 链和 β 链之间的相互作用形成了肽结合槽的裂隙。II 类 MHC 分子通过 α 链和 β 链之间的二硫键以及与 TMD 的相互作用而稳定。两条链的 TMD 都锚定在细胞的质膜上,并形成“发夹”结构,从而稳定肽结合槽。
II 类 MHC 分子的合成
II 类 MHC 分子在细胞内质网 (ER) 中合成,其过程如下:
α 链和 β 链分别合成,然后在内质网中相互结合。
这种结合是由一种称为不变链 (Ii) 的伴侣蛋白介导的。Ii 在内质网中合成,并结合到 II 类 MHC 分子的肽结合槽上,防止其与内质网中的肽结合。
这确保只有正确折叠的 II 类 MHC 分子才能被运输到细胞表面。
一旦 II 类 MHC 分子被运输到细胞表面,不变链就会被蛋白酶切割和去除,从而允许肽结合到肽结合槽上。
结合到 II 类 MHC 分子的肽来源于细胞通过内吞作用或吞噬作用内化了的细胞外蛋白。
然后将这些肽呈递给 CD4+ T 细胞,CD4+ T 细胞识别肽-MHC 复合物并启动免疫反应。
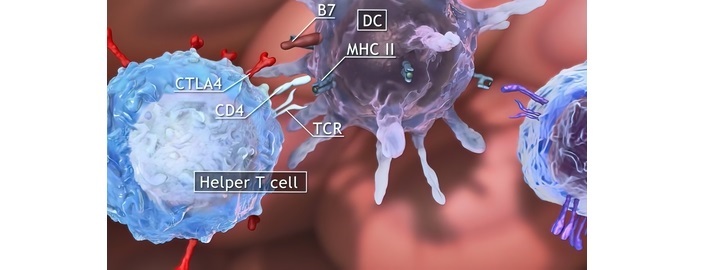
II 类 MHC 分子的意义
II 类 MHC 分子对于启动和调节适应性免疫反应至关重要。它们在将肽呈递给 CD4+ T 细胞中发挥关键作用,而 CD4+ T 细胞对于对病原体产生有效的免疫反应是必要的。
II 类 MHC 分子的意义可以概括为以下几点:
抗原呈递
II 类 MHC 分子对于将外源性抗原呈递给 CD4+ T 细胞至关重要。
抗原呈递细胞 (APC),例如树突状细胞、巨噬细胞和 B 细胞,在其表面表达 II 类 MHC 分子。
这些分子将来自细胞外蛋白的肽呈递给 CD4+ T 细胞,CD4+ T 细胞通过其 T 细胞受体 (TCR) 识别肽-MHC 复合物。
这种相互作用导致 CD4+ T 细胞的激活和分化,随后激活其他免疫细胞,例如 B 细胞、CD8+ T 细胞和自然杀伤细胞。
肽结合特异性
II 类 MHC 分子的肽结合槽对某些氨基酸具有高度特异性,允许结合多种肽。这种特异性确保只有来自外来或致病蛋白的肽被呈递给 CD4+ T 细胞,从而对入侵的病原体产生适当的免疫反应。
自身耐受
II 类 MHC 分子还在自身耐受中发挥作用。胸腺中的发育中的 T 细胞暴露于由 II 类 MHC 分子呈递的自身肽。这种暴露导致识别自身抗原的 T 细胞的删除,从而防止自身免疫反应。
移植
在移植中,供体和受体 II 类 MHC 分子之间的不相容性会导致移植组织的排斥。受体免疫系统将供体 II 类 MHC 分子识别为外来物质,导致 CD4+ T 细胞的激活和随后对移植组织的排斥。这种现象被称为同种异体反应性,是成功移植的主要障碍。
自身免疫
在某些自身免疫性疾病(如多发性硬化症和 1 型糖尿病)中,CD4+ T 细胞识别由 II 类 MHC 分子呈递的自身抗原。这种识别导致自身反应性 T 细胞的激活以及随后自身组织的破坏。这样,II 类 MHC 分子参与自身免疫性疾病的发病机制。

浏览量:110
