
 数据结构
数据结构 网络
网络 关系型数据库管理系统 (RDBMS)
关系型数据库管理系统 (RDBMS) 操作系统
操作系统 Java
Java iOS
iOS HTML
HTML CSS
CSS Android
Android Python
Python C语言编程
C语言编程 C++
C++ C#
C# MongoDB
MongoDB MySQL
MySQL Javascript
Javascript PHP
PHP植物生长调节剂及其作用机制
引言
植物生长调节剂,也称为植物激素,是调节植物生长的物质。这些PGRs充当信号分子。这些PGRs在植物体内产生的浓度极低。
这些激素由植物细胞产生,它们不仅调节植物的生长,还决定植物其他部分的形成,例如花、叶、落叶、发育和果实成熟等。简而言之,PGRs控制着植物内的所有生理过程。

植物生长调节剂的类型及其作用机制
以下是植物产生的主要PGRs。
生长素
生长素具有氨基酸前体色氨酸。它是一种促进生长的激素,它促进细胞壁松弛,增加细胞壁的塑性,并有助于细胞分裂和扩张。
它主要作用于促进根系生长。NAA防止叶片和果实过早脱落。它导致顶端优势,因此主枝的生长比侧枝快。去除顶芽会导致侧枝生长增强。它促进单性结实,从而产生无籽果实。例如:无籽草莓和葡萄。
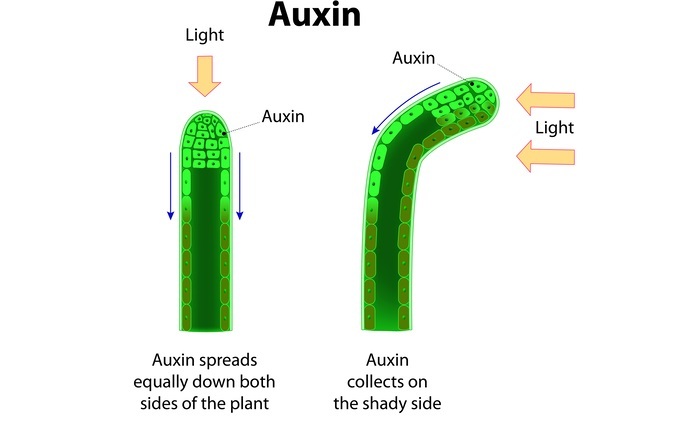
作用机制
关键参与者:生长素作为配体,TIR1(运输抑制剂反应1)是生长素激素受体。TIR1与其他F-box蛋白(如与SKP蛋白和Cullin蛋白相关的SCF)一起工作。TIR1蛋白作为E3连接酶,可以将多聚泛素连接到靶蛋白上以使其降解。
ARF生长素反应因子是一种转录因子,它有助于转录产生对抗生长素反应所需的基因。AUX/IAA是ARF的抑制剂。26S是一个蛋白水解复合体。
对于每条信号通路,都会发生转录因子的激活,而转录因子会激活某些反应基因。
在细胞核中,我们有调节区和结构基因,它们将被转录成靶mRNA,后者将用于产生蛋白质。
当植物没有接受生长素时,ARF被AUX/IAA抑制剂抑制,并且不结合到调节区。因此,不可能进行结构基因转录。
当存在生长素时,它首先被运输到植物细胞并与其细胞质中的受体结合。这种结合激活F-Box复合物,导致TIR1蛋白被激活并开始将多个多聚泛素添加到AUX/IAA抑制剂中,这导致其通过26S蛋白水解复合物降解。
然后,作为转录因子的ARF能够自由地结合到细胞核中的调节区,从而将结构基因转录成特定的mRNA,然后在细胞质中翻译成蛋白质。这些蛋白质对于细胞生长是必需的。
赤霉素 (GA)
它具有异戊二烯化合物前体IPP。GA最重要的作用是节间和枝条伸长。这使其适合增加甘蔗的产量。GA阻止种子和芽休眠并刺激其生长。

作用机制
GID1是赤霉素受体,是细胞核和细胞质中的可溶性蛋白质。首先,由种子产生的赤霉素激素与GID1受体结合,并增强GID1和DELLA蛋白(抑制植物生长的阻遏物)的相互作用。
这形成了GA-GID1-DELLA蛋白复合物,导致delta链结构,允许称为Prefoldings的蛋白质结合,然后这有助于降解这种蛋白质。一种称为PIF的转录因子与该复合物结合并转录植物DNA到mRNA,从而表达赤霉素反应基因。
然后,这种mRNA进入核糖体,在那里被翻译成蛋白质,通常是一种称为淀粉酶的酶。这种酶然后分解淀粉成糖,这是一种细胞或种子可以用来激发植物生长和伸长的能量。
细胞分裂素
它具有异戊二烯化合物前体IPP。与生长素相反,细胞分裂素阻止顶端优势并促进侧枝生长。它诱导细胞分裂并延迟衰老。最活跃和天然存在的细胞分裂素是玉米素。
作用机制
这条途径是一种双组分调控系统途径。这里,细胞分裂素是配体,CRE-1(细胞分裂素反应-1)、AHK、AHK-2、AHK-3(拟南芥组氨酸激酶)是受体。这些受体具有CHASE结构域,细胞分裂素与该结构域结合。AHP(拟南芥组氨酸磷酸转移蛋白)和ARR(拟南芥反应调节因子)。
细胞膜具有2个CRE-1或AHK受体单体。每个受体都有外侧CHASE结构域和内侧组氨酸激酶和受体结构域。现在细胞分裂素来并结合到CHASE结构域的受体上以驱动该途径。
这种配体结合诱导CRE-1或AHK受体的二聚化,从而激活组氨酸激酶。激活的组氨酸激酶作用于ATP并从中获得磷酸基团。该磷酸基团然后转移到受体的受体结构域。然后将磷酸基团转移到细胞质中无活性的AHP蛋白。
然后,AHP移动到细胞核中,并将磷酸基团转移到B型ARR的天冬氨酸残基。这然后调节A型ARR产生的mRNA的DNA和转录,后者随后翻译并产生A型ARR蛋白。这种A型ARR从AHP接收磷酸基团。
这种ARR是调节细胞分裂素反应基因的一种。这种A型ARR与B型ARR形成负反馈环,从而抑制
乙烯
乙烯也来源于氨基酸前体蛋氨酸。它增加呼吸速率,因其在果实成熟中的作用而闻名,并促进衰老和根毛形成。它导致植物的侧向生长。

作用机制
由于乙烯是负调控因子,因此它的缺失驱动该途径,而它的存在会阻止该途径。
植物具有各种乙烯受体,它们存在于内质网中。所有这些在结构上都与二聚体多通道蛋白相关。这些受体在其乙烯结合结构域中具有铜离子。这些受体还具有与蛋白CTR-1相互作用的结构域。
在没有激素的情况下,乙烯受体是活跃的,这保持CTR-1活跃。活跃的CTR-1具有活跃的丝氨酸/苏氨酸激酶结构域。CTR-1在乙烯信号传导中的功能取决于该激酶结构域的活性及其与乙烯受体的N端结构域的关联。
然后,活跃的CTR-1经历未知的信号机制,结果EIN-3被泛素化并在蛋白酶体中降解。由于这种降解,不能形成乙烯反应基因的转录。
在存在乙烯的情况下,它通过细胞壁扩散到细胞中,并进入内质网,在这里它在内质网腔中与受体结合。这种结合使受体失活,结果发生构象变化,CTR-1无法结合到该受体。因此,CTR-1失活,来自它的下游信号通路被阻断。在这种情况下,EIN蛋白不会被泛素化和降解。现在EIN-3被转移到细胞核中,并结合并激活乙烯靶基因的转录。
脱落酸 (ABA)
ABA具有异戊二烯化合物前体IPP。它具有生长抑制作用,其中它抑制芽和枝条生长,并且通常充当赤霉素的拮抗剂。在各种非生物和生物胁迫条件下,ABA水平升高,这赋予植物胁迫耐受性。它还会导致气孔关闭。

作用机制
ABA是一种干旱激素,调节气孔的开闭。气孔具有2个保卫细胞和中央的一个水孔。植物需要在白天打开气孔以获取CO2和水。
在每种气候条件下,植物都需要关闭气孔以避免水分蒸发。因此,在阳光照射下,ABA不起作用,因此气孔打开。保卫细胞具有钙储存液泡,这些液泡具有不同的钾、钙和氯离子通道。
此外,保卫细胞具有钾(钾离子的流入和流出)、氯(氯离子的输出)离子通道和质子泵。ABAR是位于保卫细胞膜上的受体,当存在配体ABA时,信号传导开始。
脱落酸 (ABA) 与 ABA 受体 (ABAR) 相互作用,导致磷脂酶 C (PLC,细胞膜相关组分) 产生 IP3。IP3 与液泡中的钙通道相互作用,允许钙离子运输到保卫细胞的细胞质中,导致钙离子内流(主要钙离子内流)。ABA 激活后还会激活活性氧 (ROS),这通过钙离子通道引起钙离子内流(次要钙离子内流)。这两种钙离子内流导致保卫细胞内钙离子浓度升高。
一方面,这些钙离子与钾离子内流通道结合并将其阻断。另一方面,这些钙离子与氯离子外排通道结合并允许氯离子外排。这些钙离子还会与质子泵结合并抑制其功能,导致氢离子浓度升高,使保卫细胞内部酸化。
因此,钾离子外排通道被激活,并通过液泡中氯离子和钙离子的外流来平衡。由于钾离子和氯离子快速移出保卫细胞,从而产生渗透压不平衡。因此,水从保卫细胞中移出,导致气孔关闭。
在阳光照射下,质子泵被激活,它具有 ATP 合酶活性并将氢离子运输到保卫细胞外部。这导致渗透压不平衡。然后它激活钾离子内流载体,然后植物产生的糖通过糖转运蛋白进入保卫细胞。由于这种不平衡,水涌入保卫细胞。这导致气孔开放。
结论
植物生长调节剂也被称为植物激素或植物内源激素。这些是在植物中天然产生的有机物质,参与控制植物的生长和许多其他生理过程,即使含量极少,也能从植物的一部分移动到另一部分。它们在植物体内起着化学信使的作用,影响植物发育的许多模式。

浏览量:597
