
 数据结构
数据结构 网络
网络 关系数据库管理系统 (RDBMS)
关系数据库管理系统 (RDBMS) 操作系统
操作系统 Java
Java iOS
iOS HTML
HTML CSS
CSS Android
Android Python
Python C语言编程
C语言编程 C++
C++ C#
C# MongoDB
MongoDB MySQL
MySQL Javascript
Javascript PHP
PHP什么是移动遗传元件?
引言
在原核生物中,遗传物质通过细胞分裂(如二元分裂)从亲代传递给子代。这种通过细胞分裂的传递被称为垂直传递。现在还有一种称为水平传递的传递方式,它不涉及细胞分裂,而是借助转化、转导和接合三种机制进行遗传物质的转移。
这三种机制借助某些能够编码蛋白质和酶的DNA序列发生,这些序列被称为移动遗传元件。它们不仅存在于原核生物中,也以转座子和逆转座子的形式存在于人类中。
移动遗传元件——概述
移动遗传元件可以定义为能够从一个位置移动到另一个位置或可以在染色体上复制的DNA片段。它们可以在原核生物和真核生物中找到,构成了基因组的大部分。
已知它们对细菌的适应性有深远的影响,不仅如此,它们还有助于生物体适应新的环境,并通过导致趋异进化来产生远亲物种。
移动遗传元件是由巴巴拉·麦克林托克发现的。她在DNA双螺旋结构和遗传密码被解释之前就发现了这些元件。由于她在这一领域的贡献,她于1983年获得了诺贝尔医学奖。
移动遗传元件的类型
原核生物和真核生物的各种移动遗传元件如下:
质粒
它们是环状DNA分子,独立于宿主染色体复制。天然存在的细菌质粒大小范围从5000到400000个碱基对。它们可以通过称为转化的过程被引入细菌细胞。它们在重组DNA技术中用作载体。
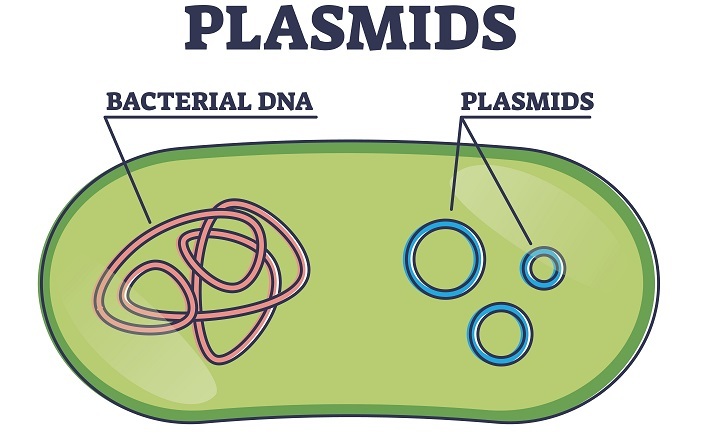
无论采用哪种方法,实际上只有少数细胞会摄取质粒DNA,因此需要一种方法来选择确实摄取了质粒的细胞。通常,宿主细胞需要质粒才能在特定条件下生长,例如赋予抗生素抗性的基因。它们也可以充当可选择标记,因为只有被重组质粒转化的细胞才能在抗生素存在下生长,使得任何含有质粒的细胞在这些生长条件下都是可选择的。
噬菌体
噬菌体具有将48,502个碱基对的DNA高效递送至细菌的机制,可用作载体来克隆稍大的DNA片段。噬菌体载体允许克隆长达23,000个碱基对的DNA片段。
转座子
这些是存在于几乎所有细胞中的DNA片段,它们从染色体上的一个位置移动或跳跃到同一染色体或不同染色体上的另一个位置。这种运动称为转座,通常不需要DNA序列同源性,新的位置或多或少是随机确定的。转座子插入必需基因可能会杀死细胞,因此转座受到严格调控,通常非常不频繁。
转座子也许是最简单的分子寄生虫,适应于被动地复制在宿主细胞的染色体中。在某些情况下,它们携带对宿主细胞有用的基因,因此与宿主处于一种共生关系。
逆转座子
一些来自酵母和果蝇等不同来源的特征明确的真核DNA转座子具有与逆转录病毒非常相似的结构,这些被称为逆转座子。它们编码与逆转录病毒逆转录酶同源的酶,其编码区两侧是LTR序列。它们通过RNA中间体从细胞基因组中的一个位置转座到另一个位置,利用逆转录酶制作RNA的DNA拷贝,然后将DNA整合到新的位点。
病毒
它们也像移动遗传元件一样起作用。通常,它们在宿主外部是非生命的,但在宿主内部是生命的。当它们感染宿主生物体时,它们会将基因从一个宿主转移到另一个宿主,甚至从两个不同物种的生物体转移。
移动遗传元件的功能
我们听说过细菌中正在发展的抗生素耐药性。这种抗生素耐药性的发展可以归因于抗生素耐药性基因,这些基因已被发现与移动遗传元件相关,使它们能够将这种耐药性传播到整个细菌群落。
移动遗传元件通过传播某些毒力因子(如外酶和外毒素)来帮助细菌。
在真核生物中,它们占据了基因组的大部分,它们是塑造大多数生物体基因和基因组的驱动力。
质粒和噬菌体等移动遗传元件可用作重组DNA技术和克隆程序中的工具。
结论
即使在发现DNA的双螺旋结构和遗传密码之前,人们就已经了解了很多关于移动遗传元件的知识。由于它们能够独立于细胞染色体复制,因此它们可以帮助介导细胞间或细胞内的DNA运输。移动遗传元件除了核心基因外,还携带其他一些基因,这些基因为宿主细胞提供了选择性优势。
某些具有经济和医学重要性的细菌表型是由这些移动遗传元件编码的。水平基因转移技术和其他相关过程为基因组学领域提供了巨大的机遇。

228 次浏览
