
 数据结构
数据结构 网络
网络 关系型数据库管理系统 (RDBMS)
关系型数据库管理系统 (RDBMS) 操作系统
操作系统 Java
Java iOS
iOS HTML
HTML CSS
CSS Android
Android Python
Python C语言编程
C语言编程 C++
C++ C#
C# MongoDB
MongoDB MySQL
MySQL Javascript
Javascript PHP
PHP逆转录转座子及其在表观遗传调控中的作用是什么?
介绍
转座子是一种可在染色体内部从一个位置移动到另一个位置的移动遗传元件,其DNA序列具有独特的移动能力。真核生物的转座子在结构上与细菌转座子相似,并且某些转座机制也相似。然而,在其他情况下,转座机制似乎涉及RNA中间体以及某些RNA病毒的进化。
表观遗传调控可以定义为通过某种方式控制特定基因工作模式的现象。
逆转录转座子
逆转录转座子编码一种与逆转录酶非常相似的酶,其编码区由长末端重复序列(LTR)侧翼。它们借助RNA中间体,利用逆转录酶将RNA复制成DNA,然后将DNA连接到靶位点,从而在细胞基因组中从一个位置跳到另一个位置。
这种转座方法在真核生物中很常见,与细菌转座不同,细菌转座是DNA直接连接到染色体内或另一个染色体的靶位点。
逆转录转座子不包含编码病毒衣壳的env基因,因此它们不能形成病毒颗粒。这使我们能够比较逆转录酶和真核转座子,这表明逆转录酶在多细胞生物出现之前就已存在。
逆转录转座子的类型
根据长末端重复序列,逆转录转座子有两种类型:
LTR逆转录转座子
它们包含一个中央编码区,该编码区由位于同一方向的LTR侧翼,这构成了所有LTR逆转录转座子的基本结构。
与其他转座子一样,它们在其几百个核苷酸对长的序列的两侧都包含短的倒置重复序列。
由于LTR序列和类似逆转录病毒的特性,它们被称为LTR转座子。
它们利用逆转录酶介导从RNA合成双链DNA,然后将其转座到靶位点并连接到靶位点。
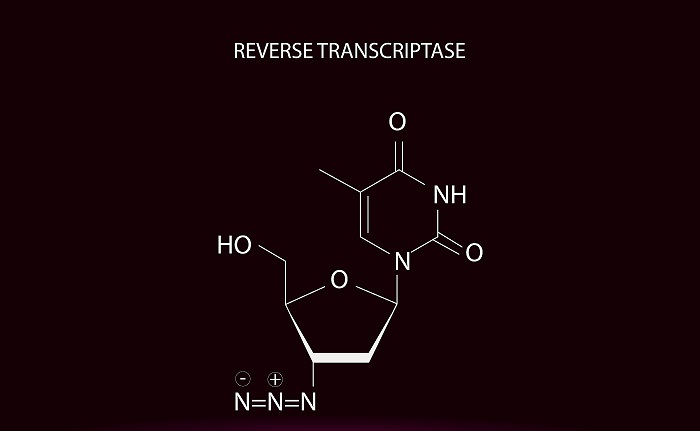
图:逆转录酶
在酵母中发现的最常见的LTR逆转录转座子是Ty元件。它们长5.9千碱基对,其中LTR的长度在两端约为340碱基对。酵母中大约有35个Ty元件的拷贝。与逆转录病毒的gag和pol基因一样,酵母Ty元件包含TyA和TyB基因。
非LTR逆转录转座子
它们是真核生物中最常见的逆转录转座子。它们也被称为反转录转座子,其两端不包含长末端重复序列,但包含同源的A和T碱基对序列以及一个末端,它是poly-A尾的修饰。
转座过程涉及形成RNA中间体,然后由元件本身编码的逆转录酶将其转化为双链DNA,然后转座到靶位点。
这些逆转录转座子最常见的例子是人类转座子,即长散布核元件(LINE)和短散布核元件(SINE)。
长散布核元件(LINE)-属于此类的人类转座元件是L1反转录转座子,其长度约为6kb,具有RNA聚合酶II识别的内部启动子序列和两个ORF。
短散布核元件(SINE)-它们长400个碱基对,包含内部启动子序列,但不编码任何蛋白质。它们是人类中第二丰富的转座元件。它们不包含任何末端重复序列,而是在一端具有A和T序列。SINE元件的一个常见例子是活跃的Alu序列。
逆转录转座子在表观遗传调控中的作用
转座子构成人类基因组的很大一部分,它们能够在染色体中从一个位点跳到另一个位点,有时它们的插入会对功能基因造成严重的损害。尽管具有某些功能,大多数这些转座元件通过甲基化或组蛋白修饰被沉默,以避免对功能基因产生任何有害影响。
但有时,这些元件会逃脱这两个过程,并导致基因功能发生变化,这个过程被称为表观遗传进化。它们的逃逸使这些位点容易受到环境刺激的影响,例如接触毒素和不健康的饮食,它们充当个体间表观遗传变异的基因座。
例子
逆转录转座子对表观遗传调控影响最常见的例子之一是Agouti可存活黄色小鼠。在这些小鼠中发现的逆转录转座子是胞浆内A颗粒(IAP),它对DNA甲基化有更深远的影响,导致个体间表型变异,例如毛色等。然而,详细的机制仍有待阐明和研究。
结论
I类转座元件也称为逆转录转座子或具有RNA中间体的转座子。这些元件在基因组的各个位点插入自身,方法是借助逆转录酶将单链RNA转化为双链DNA。它们比DNA转座子的拷贝数增加得更快。
它们也被认为参与基因的表观遗传调控以及它们赋予的不同特性,这在Agouti小鼠品种中通过逃避免基化和组蛋白修饰过程而观察到。逆转录转座子是分子生物学中研究最少的课题之一;更深入的研究可以揭示逆转录转座子在不同生物体中的其他影响。

88 次浏览
