
 数据结构
数据结构 网络
网络 关系数据库管理系统
关系数据库管理系统 操作系统
操作系统 Java
Java iOS
iOS HTML
HTML CSS
CSS Android
Android Python
Python C 语言编程
C 语言编程 C++
C++ C#
C# MongoDB
MongoDB MySQL
MySQL Javascript
Javascript PHP
PHP中枢耐受或阴性选择:功能、机制和相关疾病
简介
我们的免疫系统产生几乎无限数量的淋巴细胞,并且这些淋巴细胞中的每一个在其细胞膜上表达独特的抗原特异性受体。这些受体是由预先存在的基因片段的随机重排编码的。
抗原特异性受体通常在淋巴细胞的早期发育阶段产生,早于与特定抗原的相遇。T 细胞上的这些抗原特异性受体称为 TCR,B 细胞上的称为 BCR。
当这些淋巴细胞受体遇到特定抗原时,会受到刺激并进行增殖以产生特定淋巴细胞的克隆,最后这些淋巴细胞分化为效应细胞和记忆细胞。
免疫系统还会产生能够识别自身抗原的淋巴细胞,这些淋巴细胞称为自身反应性或自身反应性淋巴细胞。我们的免疫系统必须确保只有能够识别外来抗原的淋巴细胞才能成熟,因为自身反应性淋巴细胞的成熟会导致宿主组织损伤。
免疫耐受是我们的免疫系统耐受或不响应自身抗原以及仅针对外来抗原产生 T 细胞和抗体的能力。
免疫耐受的类型
对自身抗原或自身耐受的免疫反应通过作用于免疫系统细胞的各种机制和过程在人体内得到预防。自身耐受的建立通过一个两步过程实现,主要分为两种类型。它们是中枢耐受和外周耐受。
中枢耐受
它是检测和去除在初级淋巴器官(胸腺和骨髓)中识别自身抗原的淋巴细胞,淋巴器官发育发生在这些器官中,并且也是 T 和 B 淋巴细胞首次表达其抗原受体的部位。淋巴器官为淋巴细胞成熟提供生长因子和其他分子信号。
中枢耐受主要发生在淋巴细胞的早期发育阶段。中枢耐受的机制和过程消除了大多数自身反应性 B 和 T 细胞。但中枢耐受并非万无一失。这就是外周耐受发挥作用的地方。
T 细胞的中枢耐受或识别自身抗原(自身反应性 T 细胞)的 T 淋巴细胞的去除发生在胸腺中,而 B 细胞的中枢耐受(自身反应性 B 细胞的去除)发生在骨髓中。
大多数 T 细胞起源于骨髓并在胸腺中成熟以进一步发育。在胸腺中,T 细胞排列其受体基因,这是胸腺的重要功能。在胸腺中发育的 T 细胞称为胸腺细胞。
这些胸腺细胞中的每一个都具有随机产生的特异性。T 细胞将肽抗原识别为肽 MHC 复合物,这些复合物是我们自身的体分子,不识别氨基分离。因此,这些 MHC 称为自身 MHC 分子。
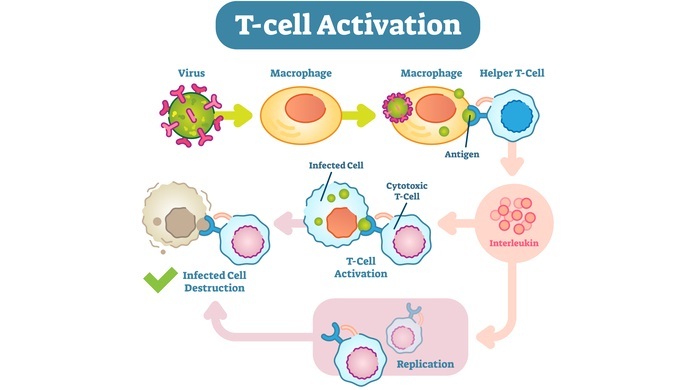
中枢 T 细胞耐受的机制
当前体 T 细胞首次从骨髓进入胸腺时,它们缺乏大多数不成熟 T 细胞特征的表面分子,并且受体基因也没有重排。后来,胸腺 T 细胞发育开始并开始表达具有随机抗原特异性的 TCR。在胸腺成熟过程中,不成熟的 T 细胞经历了一个精细的筛选过程,该过程涉及三个主要的选择事件。
非选择。
阳性选择。
阴性选择。
在胸腺内,所有有核细胞都表达 MHC I 分子,在正常情况下,这些 MHC I 分子显示自身抗原。胸腺上皮细胞也表达 MHC I 分子,并且在胸腺中,骨髓来源的树突状细胞和巨噬细胞(即抗原呈递细胞)以 MHC II 分子复合物的形式呈递肽。
这些肽抗原来源于这些细胞产生的所有蛋白质和细胞外液中产生的蛋白质。因此,在胸腺中,所有不成熟 T 细胞的命运取决于它们与自身肽-自身 MHC 分子接触的强度。

巨噬细胞与 T 细胞结合
如果不成熟的 T 细胞未能识别这些复合物,它们就会发生凋亡(T 细胞死亡),并且其中大多数具有无法识别 MHC 分子的非功能性 TCR。这种选择事件称为非选择。
如果不成熟的 T 细胞成功识别并结合到特定的肽-MHC 抗原复合物,则存活信号会传递到 T 细胞的细胞核。然后,这些 T 淋巴细胞进一步分裂和生长。此过程称为阳性选择。此处 T 细胞结合肽-MHC 复合物的程度适中。在此,T 细胞学会专注于自身 MHC 分子。

那些与肽-自身 MHC I 复合物结合非常强的未成熟 T 细胞将发生凋亡或变成 T 调节细胞。此类 T 细胞可能是低反应性细胞。这种去除选定 T 细胞的过程称为阴性选择。死亡细胞被胸腺中的巨噬细胞吞噬。
因此,由此我们可以得出结论,不成熟的 T 细胞通过中枢 T 细胞耐受的阳性选择存活下来。这些细胞具有识别 MHC 复合物中非自身抗原的潜力。此类 T 细胞离开胸腺并迁移到次级淋巴器官。
阴性选择去除所有识别自身抗原的潜在改变的 T 细胞。因此,阴性选择可预防自身免疫性疾病。并非所有自身抗原都表达在胸腺中。有些出现在其他阶段或其他组织中。此类自身反应性 T 细胞逃避中枢耐受并迁移到外周淋巴器官,在外周耐受阻止其成熟并将其消除。
胸腺细胞如何显示自身抗原?
一种称为自身免疫调节因子 (AIRE) 的转录因子导致胸腺中自身蛋白的增强和随机合成。
这导致胸腺细胞显示通常存在于胸腺外部的蛋白质的肽,例如胰腺、肺、唾液腺等。
编码 AIRE 蛋白的基因发生突变会导致自身免疫性内分泌疾病,其中免疫系统由于丰富的自身反应性 T 细胞而错误地攻击多个器官,并且阴性选择受损。
例如:自身免疫性多内分泌腺病-黏膜念珠菌病-外胚层发育不良(APECED),其特征是复发性念珠菌感染、甲状旁腺功能减退和肾上腺功能衰竭。

人甲状旁腺激素
B 细胞的中枢耐受
基质细胞是骨髓内 B 细胞中枢耐受阴性选择的主要部位。与 T 细胞相比,B 细胞的耐受性不完整。这些基质细胞具有其自身抗原,有助于 B 细胞的选择。
大多数这些抗原是多价的,可有效交联 BCR。如果一些具有其特定 B 细胞受体或 BCR 的 B 细胞选择非常强烈地粘附到这些基质细胞的抗原上,那么它们将保留在骨髓中,并且可以具有以下 3 种可能的诱导耐受的机制结果。
受体编辑 - 重排 B 细胞受体,使其不与自身抗原反应。
克隆消除 - 通过凋亡杀死
无能状态 - 变得失活
因此,最终,自身反应性 B 细胞在此处变得无功能,因为它们可以产生导致自身免疫性疾病的自身免疫抗体。如果 B 细胞不粘附于基质自身抗原,则它们会迁移到外周淋巴器官,在那里它们会遇到特定抗原并被激活、增殖和分化。这些效应 B 细胞称为浆细胞,它们表达 IgD,然后进入血液。

B 细胞激活
结论
人体中的每个淋巴细胞都表达抗原特异性受体。用于自身和外来抗原。我们的免疫系统能够对大量不同的外来抗原做出反应,同时对自身抗原具有耐受性。
这种免疫系统对抗原的无反应状态称为免疫耐受,这种对自身抗原的无反应状态称为自身耐受。如果免疫系统无法实现这种自身耐受,就会导致自身免疫性疾病,其中免疫系统开始攻击自身组织。
自身免疫性疾病是此类后果的最终结果,其中个体的自身组织和器官受到自身免疫系统的攻击。自身耐受分两个步骤完成,分别是中枢耐受和外周耐受。

94 次浏览
