
 数据结构
数据结构 网络
网络 关系型数据库管理系统
关系型数据库管理系统 操作系统
操作系统 Java
Java iOS
iOS HTML
HTML CSS
CSS Android
Android Python
Python C 编程
C 编程 C++
C++ C#
C# MongoDB
MongoDB MySQL
MySQL Javascript
Javascript PHP
PHP共同进化过程与生命史理论
环境学家经常声称,我们对自然的影响与自然对我们的影响一样大。大量研究突出了这两个实体之间相互作用。然而,我们无法理解它们的活力或两者之间的联系。然而,我们不应该对该理论证明这一点感到惊讶。支持更大大脑的环境变量也有利于更多地投资于保护,因为大脑袋的好处是长远的。

另一方面,减少死亡率的外源性环境过程促进了生存成本的增加,因此促进了对智力资本的更大投资。在哺乳动物中,预期寿命和脑容量呈正相关。
共同进化过程与生命史理论
传统的生命史理论 (LHT) 程序无法准确地模拟这种共同进化。它们假设死亡率存在一个不受选择影响的“外在”组成部分,这使得它们在解释其他生命史变量(如初次生育期和衰老速度)方面具有优势。然而,这种方法在理论上是不够的,因为生物体对其几乎所有死亡原因都具有一定的控制权(例如,通过修改旅行习惯以逃避迫害或增强免疫系统)。它也存在分析上的局限性,无法理解死亡率是如何完全变化的。
一种更有效的方法是假设,随着环境条件的变化而变化的是死亡与减少死亡所付出的努力之间的操作联系,而不是固定的死亡率。一种概念化外源性变异性的方法是将其视为不同的“攻击”类型和强度。例如,温暖潮湿的气候有利于病原体的生长,从而增加了危害生物的疾病的频率和种类。此外,改善这些状况的努力与死亡率下降之间存在联系。
行为生态学:成本效益分析与生命史理论
LHT 是优化方法的一部分,该方法试图通过权衡某个领域内潜在策略的成本和收益,来确定在缺乏遗传或发育限制的情况下,生物进化中可能出现的策略。这种方法更广泛地应用于行为生态学和理论生物学。
在 60 年代和 70 年代,这种方法改变了理论生物学。在此之前,科学家们并没有始终如一地将明确的经济因素纳入选择中。除了生命史理论之外,这还导致了大量新假设的出现,包括进化心理学家所依赖的许多“进化中间理论”,如父母投资理论、亲子冲突、性别分配理论等。
成本效益分析现在是进化生物学的基础,也是行为生态学中首要的方法。LHT 对于成本效益分析来说并不是必要的。例如,觅食策略可以根据储存能量的益处和能量消耗的成本进行建模。最佳方法是最大化瞬时净卡路里消耗量。由于这种建模没有考虑策略选择对时间的跨期影响,因此它不是 LHT。当建模明确考虑可能的方法对生物体将来可能达到的所有年龄段的适应度结果的影响时,则称其采用了生命史方法。
LHT 最初侧重于生活经历的顺序。然而,生物学家发现,需要清晰的生命史视角来理解通常不被认为是生命史事件的现象。因此,在许多领域,LHT 逐渐取代了成本效益分析。LHT 是一种理解选择的通用分析技术,而不是它解释的现象。
诚实信号理论
信号理论的最新进展证明了这一想法。“诚实”的质量信号是指高质量个体(“大型信号发送者”)能够负担得起而低质量个体负担不起的信号。传统上,人们认为这些信号是活力标志,主要信号发送者比其他人更有可能存活。从理论上讲,他们可以“花费”比其他人更多的生存能力来发出信号,从而通过提高生育能力来提高适应性。免疫能力信号模型是这一概念的一个显著例子。
人们被认为在寄生虫抵抗力方面存在差异,高质量的个体通过免疫抑制(例如,睾酮依赖性信号)向潜在的配偶宣传其寄生虫抵抗力。活力指标与随机信号进行了比较。后者不是质量的真正指标,因此与生存能力相关;相反,它们可能主要发展是为了增加吸引力。
Grafen 从模拟活力指标选择开始。他假设所有个体,无论质量如何,从一定量的信号中获得相同的适应度收益(即,源自信号向他人广播的交配优势,他们没有根据区分个体适应性的依据,除非通过信号)。当与发展和维持特定信号水平相关的适应度成本(以死亡率为货币)对于高质量个体低于低质量个体时,信号可以进化到显示质量(即,它由于质量函数的差异成本而进化,而不是差异收益)。
信号“诚实”地传递质量,因为低质量个体“欺骗”并产生更大的信号并非符合他们的最佳利益;他们将因此产生的死亡率成本将超过他们从更大的信号尺寸中获得的生育能力收益。
关于分配的选择执行
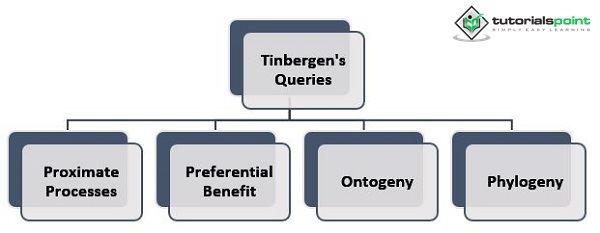
为了全面了解生命史,需要考虑廷贝根的所有问题——近端过程、偏好利益、个体发育和系统发育。理解近端机制如何发挥作用和进化至关重要。形成和执行关于一个人生命史的决策的过程是什么?此外,这些过程是如何产生的?LHT 指的是个体“选择”的分配,这意味着他们以不同的方式将时间和精力用于不同的生活活动。
LHT既不需要也不建议“适应度最大化者”或权衡成本和收益的小人。它不包含“决策者”。相反,选择可能塑造了特定的心理和生理过程,使其依赖于外部变量,这些变量以可能在过去在不同的条件和生命阶段下产生最大适应性的方式改变能量的最佳利用方式。
关于如何分配能量的决策通常需要对多个系统进行同步调整。例如,应将更多资源分配给繁殖,而将更少资源分配给生长。在总支出较少时,安排对感染的免疫反应可能是理想的。通常需要分布在多个体细胞系统上的相互作用和监督网络来实现自适应同步。实际上,内分泌系统被创造出来发挥这种作用。
内分泌系统充当内部信使。在一个部位(如肾上腺皮质或性腺)释放的激素会被其他部位(如大脑结构)的传递体“吸收”,并在那里产生影响。因此,内分泌系统能够调节能量分配,同时控制范围广泛的不同过程。当然,受体的位置及其对激素结合的反应决定了它们执行此操作的具体方式。该机制可能已通过选择进行微调,以便内分泌活动以最佳方式改变能量分配。
以影响繁殖的激素为例。青春期期间,肾上腺发育引发了男女一系列持续超过十年的发育过程。控制女性能量平衡的机制导致脂肪堆积和周期性月经。正如雌激素和其他激素控制的那样,更多能量被用于繁殖特征和活动,即额外的性特征,而发育最终会减慢。男性开始产生大量的雄激素,这会导致更多的肌肉质量和对寻求配偶活动的投资,如社交竞争力和运动能力。同时,免疫系统健康方面的一些支出被取消了。
心理机制的操控对于男女双方的同步反应网络都至关重要,如同能量消耗的改变一样。在较短的时间内,生殖激素也会控制不同的投资策略。在整个怀孕期间,母体能量必须分配给正在发育的胎儿,这通过脑、卵巢和子宫组织之间的化学途径(例如,使用促性腺激素)来实现。通过这种方法,可以终止那些“未展现”其潜力的胚胎。随着男性生育子女,他们的睾酮水平下降,使他们能够将生殖努力从交配转移到照顾子女。
许多其他内分泌或其他通讯网络调节能量输出、组织特异性吸收和认知因素,以应对历史上曾表明最佳利用方式发生变化的其他事件。例如,糖皮质激素调节压力反应,肾上腺素对战或逃反应中能量释放和利用的影响,以及其他组织对免疫功能和能量利用的调节,都是内分泌和其他通讯网络调节能量释放、组织特异性吸收的例子。
这些机制中没有任何一个需要“中央指挥中心”来协调众多受体部位的活动,并且它们的最终作用会影响其他部位。相反,这种协同作用类似于足球队进行的进攻战术,每个球员都有预定的任务,当与其他球员的任务一起执行时,旨在产生适应性的结果。选择塑造了“战术”的“设计”,其中包括赋予“球员”的角色。
结论
努力的重新分配通常涉及心理和生理过程;除非识别并响应触发它们的事件,否则重新分配无法发生。大多数情况下,很难理解起作用的心理过程。因此,实践和理解其复杂性对于获得适当的结果至关重要。

59 次浏览
