
 数据结构
数据结构 网络
网络 关系数据库管理系统 (RDBMS)
关系数据库管理系统 (RDBMS) 操作系统
操作系统 Java
Java iOS
iOS HTML
HTML CSS
CSS Android
Android Python
Python C语言编程
C语言编程 C++
C++ C#
C# MongoDB
MongoDB MySQL
MySQL Javascript
Javascript PHP
PHP植物器官分化的分子基础
引言
全能性是指细胞在适当的营养和环境条件下能够发育成一个完整的新个体的能力。孢子和受精卵都具有这种全能性。在植物中,分生组织细胞具有这种全能性。
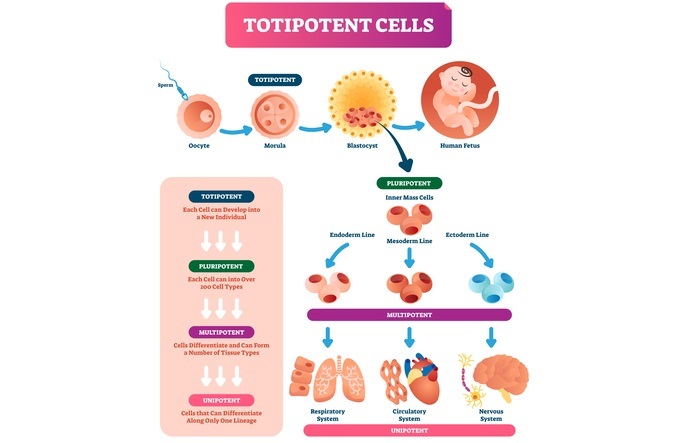
在全能性中,每个细胞都可以形成组织、器官或整株植物。由于植物的这一特性,我们可以观察到组织再生。
细胞全能性的分子基础
每个生物体都具有几乎相同的DNA,这种遗传物质在组蛋白和其他重塑复合物的帮助下被包装成染色体或染色质。这种染色质可以根据发育阶段、周围环境、压力、营养供应、激素和其他因素自由地压缩或松弛。
这些因素决定并向细胞发出信号,使其要么分裂形成一个相同的细胞,要么形成一个具有相同功能的细胞(这通常发生在有丝分裂过程中),或者这些因素可能通过改变其染色质结构来对细胞进行编程,允许一组特定的基因被激活而其他基因保持关闭,从而分化形成另一种类型的特化细胞。
这个过程被称为细胞分化。获得的分化细胞可以形成其自身类型的另一个细胞,或者进一步分化和特化其功能,或者根本不分裂。
例如,这是干细胞产生各种类型的细胞特化,最终形成一个完整的多细胞生物体的细胞分裂和分化的结果。然而,在正常情况下,分化细胞类型特化为特定命运后,不能产生另一种细胞类型。但如果像激素这样的因素得到适当的改变,分化细胞可以进行基因重编程,变得未分化,甚至重新分化,产生具有不同功能的不同细胞类型。
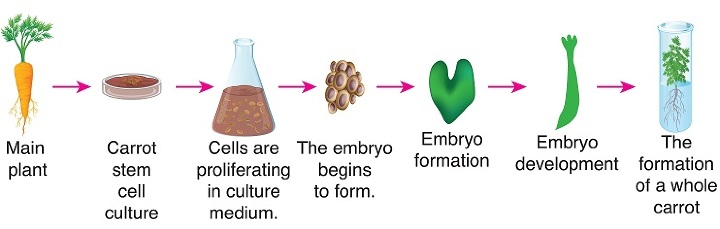
这意味着我们可以取任何活的植物细胞,提供正确的因素和条件,它就能够发育成整株植物。这是植物全能性的基础。例如:如果我们取来自胚胎的细胞,它们已经是未分化的,自然可以形成整株植物。但是如果我们取顶端分生组织,它通常具有形成芽的能力。
同样,根端分生组织只会产生根。但是我们可以处理植物并改变激素或其他因素,以使任何细胞(可能是芽、根或任何其他类型的细胞)去分化、重新分化、分裂并形成整株植物。由于全能性,利用组织培养可以快速地从植物组织中繁殖数十亿株植物。
植物的分化
在植物中,分生组织细胞被称为全能的,并且由此产生的细胞(如筛管细胞、叶细胞、根细胞等)可以分化成不同类型的细胞。源自分生组织的细胞分化并成熟以执行特定功能。细胞失去原生质,并发育出木质纤维素的次生细胞壁,形成输导组织来运输水分。
形成的叶细胞与其他细胞(如根细胞和筛管细胞)具有不同的结构和功能。叶片含有参与光合作用的叶绿体,而根则需要准备好吸收水分和养分,因为它们具有根毛。
负责分化成器官甚至整株植物的所有基因虽然存在于不同植物部分的所有细胞中,但只有特定器官或部分的特定基因表达,其余基因将处于非活动状态。只有在含有植物激素的培养基上生长的此类细胞表达所有基因,因此可用于产生所有不同的器官或整株植物。
为了表达全能细胞,首先,分化细胞必须经历去分化,然后是再分化。
去分化:成熟细胞恢复到其分生组织阶段并形成未分化愈伤组织的现象称为去分化。
再分化:未分化细胞形成整株植物或器官的能力称为再分化。例如:愈伤组织到器官。
如果我们有一个通过去分化过程诱导的全能细胞,它将产生一团称为愈伤组织的细胞,处于未分化阶段。然后,在培养基中用植物激素诱导这团细胞进行胚胎发生(胚状体的形成)或器官发生(对于愈伤组织发生,即芽的形成;对于根的形成,即根的发生;以及对于芽和根的形成,即茎根发生)或组织发生(对于木质部和韧皮部的形成,即维管组织)。
分生组织
分生组织是含有未分化细胞(称为分生组织细胞)的组织,这些细胞具有产生整株植物各个器官的能力,并负责植物的生长。通常,分化细胞不能分裂并产生其他类型的细胞。分生组织细胞具有干细胞的特性,分为三种类型
顶端分生组织。
居间分生组织。
侧生分生组织。
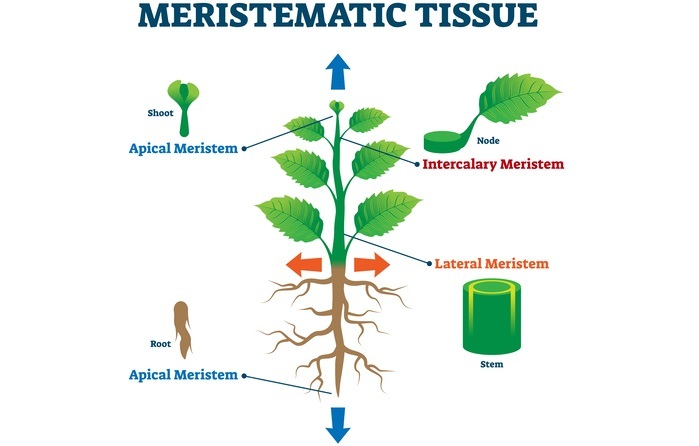
顶端分生组织位于芽或根的尖端,在那里产生生长素,从而发生初生生长,使长度增加。这种顶端分生组织又分为两种类型:SAM(芽顶端分生组织)和RAM(根顶端分生组织)。
芽顶端分生组织 (SAM)
它是开花植物胚胎发生最重要的部位。叶原基、萼片、花瓣、雄蕊和子房都起源于此。
SAM有三层L1、L2和L3。L1是外层,形成芽的表皮。L2第二层是形成植物体的组织来源,L3第三层又是形成植物体的组织来源。每一层都有自己的干细胞,所有三层都参与茎和侧器官的形成。
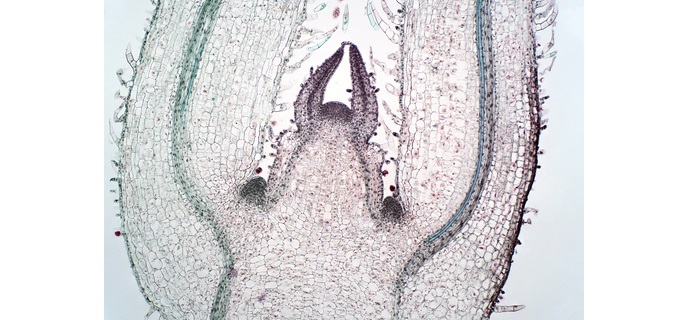
芽分生组织
SAM中主要存在四种类型的细胞群。
干细胞。
干细胞的子细胞。
组织中心。
器官起始的创始细胞。
从顶端分生组织起始芽需要许多基因的作用。主要涉及三类基因。
STM(无芽),它是KNOX家族的成员——KNOX基因在SAM中表达,对于促进SAM的身份非常重要。这些对于芽的形成是必要的,但对于根不是必要的。
WUS(Wuschel)——存在于组织中心,促进细胞分裂。
CLAVATA (CLV)——CLV1、CLV 2和CLV3,通过控制细胞分裂速率来调节SAM中干细胞库的大小。CLV1编码富含亮氨酸的受体激酶 (LRRK),一种受体,CLV3编码CLV1的配体,CLV2编码一种没有激酶结构域的受体。
STM和WUS通过使分生组织保持未分化状态来积极促进SAM的生长,因此它们可以自由地产生任何器官,如叶、茎等,而CLV基因则抑制这种生长,因为它们限制了未分化细胞的数量。CLV基因负调控WUS的表达。CLV1和2与CLV3蛋白的相互作用抑制WUS的表达,从而最小化SAM的大小。CLAVATA基因的突变导致更大的SAM。
根顶端分生组织 (RAM)
RAM是次顶端的,因为顶端有根冠。它甚至在远离轴的地方产生细胞以启动根冠。根的顶端不与侧器官的形成有关,如芽。这里的侧器官是在远离顶端分生组织的地方形成的。与芽一样,根顶端的尺寸和形状没有结构变化。
许多理论被提出用于根的组织
顶端细胞理论。
组织原理论。
Korper-Kappe 理论。
静止中心。
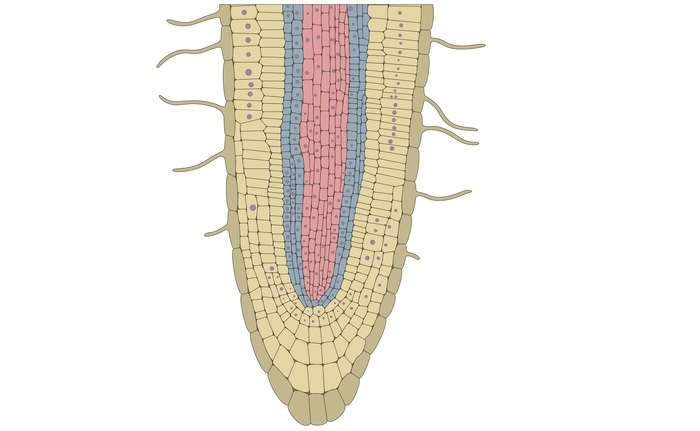
植物根系结构
结论
已经分化并丧失分裂能力的细胞,在特定条件下可以恢复分裂能力的现象称为去分化。这种分生组织分裂并产生再次丧失分裂能力并成熟以执行特定功能的细胞的过程称为再分化。
全能性是指细胞形成完整生物体或植物的全部遗传潜能,细胞全能性是分生组织细胞分化成各种器官甚至整株植物的重要概念。
细胞或组织成熟后的最终结构取决于细胞在植物中的位置。例如:远离根端分生组织的细胞分化成根冠,靠近的细胞形成表皮。

浏览量:300
