
 数据结构
数据结构 网络
网络 关系型数据库管理系统
关系型数据库管理系统 操作系统
操作系统 Java
Java iOS
iOS HTML
HTML CSS
CSS Android
Android Python
Python C 编程
C 编程 C++
C++ C#
C# MongoDB
MongoDB MySQL
MySQL Javascript
Javascript PHP
PHP蛋白质靶向或蛋白质分选
蛋白质靶向
蛋白质被传递到细胞内或细胞外的适当位置的生物学过程称为蛋白质靶向或蛋白质分选。蛋白质可以分泌到细胞外部、质膜、各种细胞内膜、细胞器的内部或质膜。
蛋白质本身包含控制传递机制的信息。正确分选对细胞至关重要,分选出现问题与许多疾病有关。

简介
在真核细胞中,各种蛋白质必须被传递到细胞内的不同位置,或者在某些情况下,被输出到细胞外环境。适当的蛋白质是如何到达相应位置的?
蛋白质通过细胞中存在的各种运输机制被传递到其目标位置,有点像分子邮政服务。在这些系统中,蛋白质使用分子标签(通常是氨基酸序列)进行“地址”标记,以便传递到特定的位置。让我们看看这些传递系统是如何工作的。
信号肽
信号肽通过充当靶向信号,使细胞运输系统能够将蛋白质引导到特定的细胞内或细胞外区域。尽管尚未发现信号肽的共有序列,但其中许多仍然具有独特的结构三部分:
靠近N端的亲水性、带正电的区域。
信号肽中一段 10 到 15 个疏水性氨基酸的区域。
靠近C端的略微极性区域,当它们位于靠近切割点时,有利于具有较短侧链的氨基酸。
蛋白质到达目的地后,信号肽通常会被信号肽酶切割。
因此,大多数成熟蛋白质中不存在信号肽。虽然过氧化物酶体的C端延伸部分存在靶向序列,但大多数信号肽位于N端。
蛋白质易位
用于分泌或特定细胞器的蛋白质必须被运输,因为核糖体将mRNA翻译成蛋白质是在细胞质中进行的。这个过程被称为共翻译易位,它也可以在翻译过程中发生,或者翻译后易位,它发生在翻译完成后。
蛋白质靶向
线粒体
大多数线粒体蛋白作为具有摄取肽信号的胞质前体产生,而细胞器中的一些蛋白质是由线粒体DNA产生的。
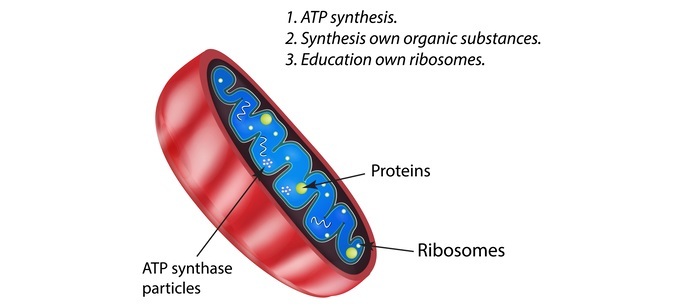
根据其序列,由胞质伴侣蛋白hsp70结合并发送到线粒体的未折叠蛋白可能定位于四个不同的位置。它们可能被引导到内膜、膜间隙、外膜或线粒体基质。健康和疾病与其中一种或多种机制的缺陷有关。
线粒体基质
当多肽到达基质时,信号序列会被加工肽酶切割,剩余的序列在等待适当折叠和作用时会被线粒体伴侣蛋白保持。
线粒体在氧化磷酸化过程中产生的电化学梯度推动并拉动多肽从胞质到膜间隙,然后到基质。
由于积极参与代谢的线粒体,膜间隙具有正电位,而基质具有负电位。靶向序列的带正电部分被基质的负电位引导到其预期位置。
线粒体内膜
根据其整体序列,线粒体蛋白可以通过三种不同的途径之一被引导到内膜,尽管它们仍然可以通过使用TOM20/22导入受体复合物和TOM40通用导入核心从外膜进入。
在靶向内膜的蛋白质的第一条途径中,其具有将多肽引导到包含先前所述易位复合物的内膜复合物的基质靶向序列,其步骤与靶向基质的蛋白质的步骤相同。
线粒体外膜
前体蛋白通过使用内部靶向序列产生跨磷脂双层的疏水性α螺旋或β桶,与外膜易位复合物相互作用以将其嵌入膜中。此过程称为外膜靶向。
这可能通过两种不同的途径发生,具体取决于前蛋白的内在序列。如果前蛋白具有能够构建α螺旋的内部疏水区域,则它将使用线粒体导入复合物并横向运输到膜中。
与β桶构建蛋白相关的并且具有疏水性内部序列的前蛋白将从上述外膜复合物TOM20/22导入到膜间隙。
当它们与TIM9/10膜间隙蛋白复合物结合时,它们会被转移到外膜的分选和组装机制(SAM),该机制将靶向蛋白作为β桶横向化。
叶绿体
根据其序列,蛋白质可以被发送到叶绿体的基质、类囊体腔、外被膜、内被膜或类囊体膜。
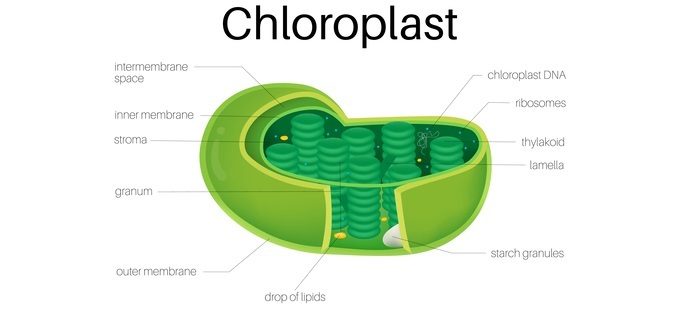
由于它们通常缺乏可切割的分选序列,因此靶向叶绿体被膜的蛋白质会被膜分选复合物横向置换。
叶绿体被膜内的Toc和Tic复合物对于从胞质中大量导入前蛋白是必需的。
叶绿体外部膜的易位酶称为Toc,叶绿体内部膜的易位酶称为Tic。Toc复合物的功能由至少三种蛋白质组成。
基质
Tic复合物中必须存在至少五种不同的Tic蛋白才能形成穿过内被膜的易位通道。到达基质后,基质导入序列会被信号肽酶切除。
根据目前的知识,ATP通过基质HSP伴侣蛋白的水解驱动这种蛋白质运输机制到基质,而不是在线粒体中产生的推动蛋白质导入的跨膜电化学梯度。
结论
双靶向肽通常具有介于两个特定肽之间的特征。这些蛋白质的靶向肽包含相对较少的带负电荷的氨基酸和大量的碱性和疏水性氨基酸。
它们包含较少的丙氨酸和更多的亮氨酸和苯丙氨酸。与线粒体和叶绿体蛋白相比,双靶向蛋白具有更疏水的靶向肽。仅根据其物理化学特性来确定肽的双靶向状态是费力的。

2K+ 次浏览
