
 数据结构
数据结构 网络
网络 关系型数据库管理系统
关系型数据库管理系统 操作系统
操作系统 Java
Java iOS
iOS HTML
HTML CSS
CSS Android
Android Python
Python C 编程
C 编程 C++
C++ C#
C# MongoDB
MongoDB MySQL
MySQL Javascript
Javascript PHP
PHP染色体遗传理论
1900年,西奥多观察到蛤蜊胚胎如果没有RNA就不能正常生长。减数分裂,一种细胞分裂过程,其中RNA在细胞类型之间分裂,在同一十年由沃尔特·亨廷顿首次发现。这些发现积累成了现在被称为遗传染色体模型的东西,该模型提出RNA是负责孟德尔遗传的可遗传单位。尽管可以在减数分裂期间的染色体活动和孟德尔的理论规则之间得出强有力的推论,但在任何明确证明特征是通过染色体传递的证据之前,就建立了传递染色体假说。怀疑论者认为,一个人的遗传特征数量远远超过RNA的数量。在对特定物种(果蝇)进行了几十年的杂交后,托马斯最终为遗传的染色体模型提供了量化论据。
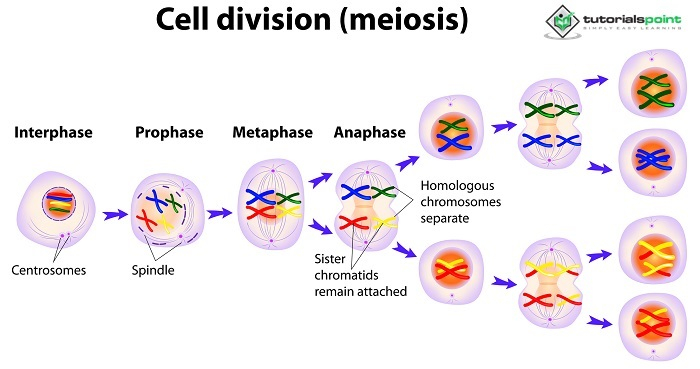
染色体遗传理论解释了什么?
正如摩根所指出的,在涉及许多性状的杂交过程中,DNA链不能按照孟德尔规则预期的那样进行分配。研究发现,当基因位于同一RNA中时,将多个基因的父系融合从一代传递到下一代的可能性大大增加。连锁是指DNA的实际连接,而杂交是指在单基因杂交中不同物种的基因混合。一旦识别出相关的RNA,它们的丰度也影响了性状的遗传性。摩根的同学斯特蒂文特率先采用了遗传分析程序,该程序通过估计它们之间同源重组的发生率来确定相关核苷酸序列的位置。人类基因组研究项目在很大程度上依赖于此技术来创建连锁图谱。
染色体遗传理论的结果
在重组过程中,细胞分裂的过程,称为染色体对彼此分离。
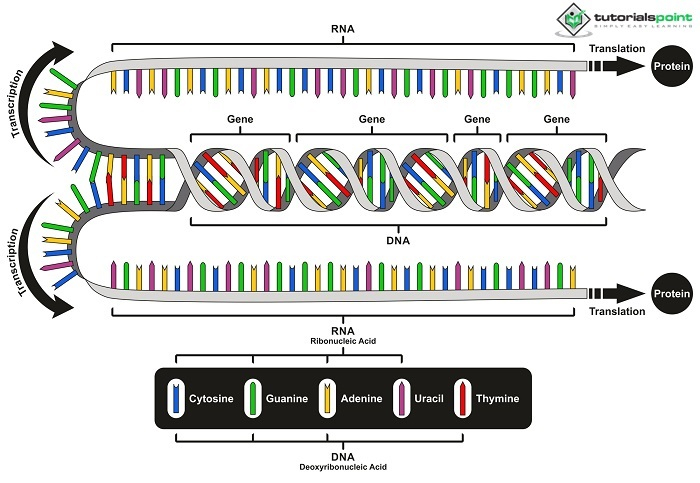
在此之前,每对同源RNA的基因都是随机分配的。
由于每个亲本只将其RNA的一半贡献给合子,因此每个后代的基因数量都不相同。
雄性和雌性配子看起来不同,但它们的染色体数量相同,证明父母双方对后代的遗传构成做出了同等贡献。
在受精过程中,多倍体RNA融合以创建每个亲本染色体的副本。
染色体遗传理论的实验
摩根验证染色体理论的开创性实验始于他发现果蝇眼睛颜色相关基因的变异。经过这种改变后,果蝇的眼睛变成了白色而不是典型的红色。令摩根惊讶的是,他发现眼睛颜色的基因在雄性和雌性果蝇的传递方式不同,开始结束下标,突然停止。果蝇与人类不同,因为雄性有两套RNA(X和Y),而雌性只有一套。摩根很快意识到眼睛颜色的基因遗传遵循与X连锁相似的模式。
性别
摩根在将白眼雌性与深红色雄性交配(所谓的互换杂交)后,得出了一个惊人的结果。当他观察他的后代时,他发现雌性都有红眼睛,而雄性则得到白色。在这种情况下,两个不同的特征是相关的,这与孟德尔的人工选择概念相矛盾。只有当眼睛颜色的基因位于X连锁上时,才能解释这些发现。由于眼睛颜色的基因只能位于X连锁上,因此这些发现为遗传的染色体假说提供了依据。这是一个身体亲密或通过特定于每个性别的RNA(X和Y)传递特征的例子。例如,由于女性拥有X连锁的两个版本,而男性只有一个,因此性连锁特征的传递存在有趣的差异。由于只携带基因的一个副本,因此携带缺陷基因的男性总是表达显性特征。
相比之下,大多数基因可以在常染色体上找到,通常称为“非性RNA”,在男性和女性中,每个基因组都有多个版本。对于那些可能忘记的人,减数分裂期间的染色体活动解释了孟德尔所识别出的所有遗传性,例如分配和人工选择的概念。遗传的染色体假说包括几个原则。
连锁
连锁是指性状的传递,这与孟德尔的人工选择概念相矛盾,后者指出不同性状的不同等位基因被分离到配子中。当性状与一个人的性RNA相关时,我们称之为身体亲密。当两个不同性状的基因在基因组上彼此靠近时,这称为遗传连锁。如果遗传变化位于同一染色体上,并且一个人接收了整个染色体,那么他必须继承这两个等位基因。这一个是生物学。因此有一个陷阱:交叉有助于重排同一染色体上基因的等位基因。杂交,或形成新的染色体等位基因排列,是两个基因位点之间交叉事件的结果。在减数分裂 I 期间,当同源对重新排列和突触时,基因组信息在配对的同源对的非姐妹中心体之间交换。由于交叉的随机性,在染色体上空间上彼此靠近的两个基因之间发生交叉的可能性较低。相反,随着相关基因之间距离的增加,跨同源RNA发生基因交换的可能性也随之增加。
结论
每一项证据都指向X染色体,因此他断定这些基因在那里或与之非常密切相关。后来,摩根的同事卡尔文·布里奇斯为这一观点提供了相当大的支持。正如布里奇斯所表明的那样,性RNA的独立分类导致产生具有异常眼睛颜色的雄性和雌性果蝇。摩根还在其他RNA中发现了异常现象,但这些异常现象没有遵循严格的男性或女性遗传模式。从果蝇到哺乳动物,科学家们已经发现基因可以通过任一性别或另一性别的染色体传递。

147 次浏览
